Interaksjon mellom lys og CO2 fremmer veksten hos vannplanter
|
Interaksjon mellom lys og CO2 fremmer veksten hos vannplanter - del 1 © www.tropica.com | |||
| At kun én ressurs om gangen begrenser veksten hos planter, er en teori, som fortsatt forekommer selv i nyere lærebøker. Denne teorien er også bedre kjent som Liebigs prinsipp. Justus Liebig var en tysk kjemiker, som blant annet arbeidet med næringsstoffbehov hos landbruksavlinger. Han forfektet, at én og kun én faktor, kunne begrense planteveksten til en gitt tid. Det er usikkert, hvorvidt Liebig selv har forfattet en tilsvarende illustrasjon som den på figur 1, men den delvis fylte baljen er simpel og intuitiv, og den har formodentlig i høy grad medvirket til å bibeholde Liebigs prinsipp som en allment akseptert modell for ressursbegrensning. Det mangelfulle ved dette enkle prinsippet har vært kjent for terrestriske planter gjennom flere årtier og innenfor de siste 20 år har også akvatiske plantefysiologer erkjent, at flere ressurser samtidig kan begrense vekstprosessene i plantene. Få studier på akvatiske planter har vist, at interaktive effekter av lys og CO2 i fotosyntese kan resultere i egentlig vekst (Maberly 1985, Madsen og Sand-Jensen 1994). Her viser vi resultater fra eksperimenter, hvor vi har skapt samtidig begrensning av lys og CO2, som begge er viktige ressurser, som vanligvis er begrensende for planters vekst i naturen. | |||
|
Figur 1. Figuren viser den klassiske illustrering av Liebigs prinsipp. I denne situasjonen er elementet Bor begrensende for planteproduksjonen, og det illustreres ved, at vannet renner ut av baljen når ressursbegrensningen er nådd. | ||
|
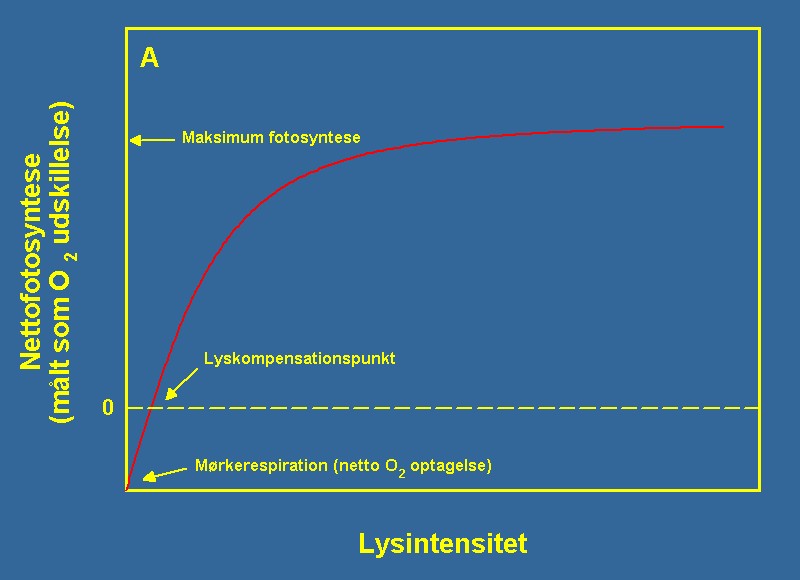
Den generelle sammenheng i mellom lys og fotosyntese hos en vannplante er illustrert i figur 2A. Ved lav lysintensitet er fotonmengden for lav til å opprettholde av en positiv fotosyntese og nettoproduksjonen av oksygen hos planten er negativ, dvs. at de respiratoriske prosesser forbruker mer O2 enn det produseres ved fotosyntesen. Den lysintensitet, hvor fotosyntesen er lik respirasjonen, defineres som plantens lyskompensasjonspunkt. Ved stadig stigende lysintensitet økes fotosyntesen fortsatt lineært med lyset, men ved enda høyere lysintensitet blir O2-produksjonen proporsjonalt mindre inntil planten når sin maksimale fotosynteserate. Ytterligere økning av lyset vil fra dette punkt ikke øke fotosynteseraten. I naturen er lyset ofte begrensende for vannplanters vekst. Vannet absorberer i seg selv lyset effektivt og omdanner energien til varme. Oppløste organiske stoffer - eksempelvis de brunfargede humussyrene - absorberer likeledes effektivt lyset i vannsøylen. Vannets lysabsorbering samt innholdet av organiske stoffer bestemmer dybdegrensen for vannplanters vekst i naturen. Til tider blir lystransmisjonen for lav til at undervannsplanter kan gro og kun flytebladsformer samt egentlige undervannsplanter kan trives. Evolusjonen har, som følge av lysets betydning for veksten hos planter, utviklet et meget effektivt system for lysutnyttelse hos undervannsplanter. Hvis plantene har høy tilgjengelighet av næringsstoffer, investerer de generelt mer energi i lysabsorberende pigmenter så som karotenoider, xanthophyller og i særdeleshet klorofyll. Klorofyll er det grønne pigmentet, som absorberer energien og overfører det til kjemisk energi til bruk for vekst i cellene. På denne måten sikrer plantene, at den mengde lys, som når plantens overflate, absorberes og brukes til energetiske formål fremfor bare å bli transmittert gjennom plantevevet. Det er likeledes nødvendig med en stor pulje av klorofyll for å opprettholde en høy maksimal fotosyntese - men en stor pulje av klorofyll er ikke mye verdt for planten, hvis den absorberende energi ikke kan avsettes til de energikrevende prosesser, som assimilerer uorganisk kullstoff til kullhydrater. | |||
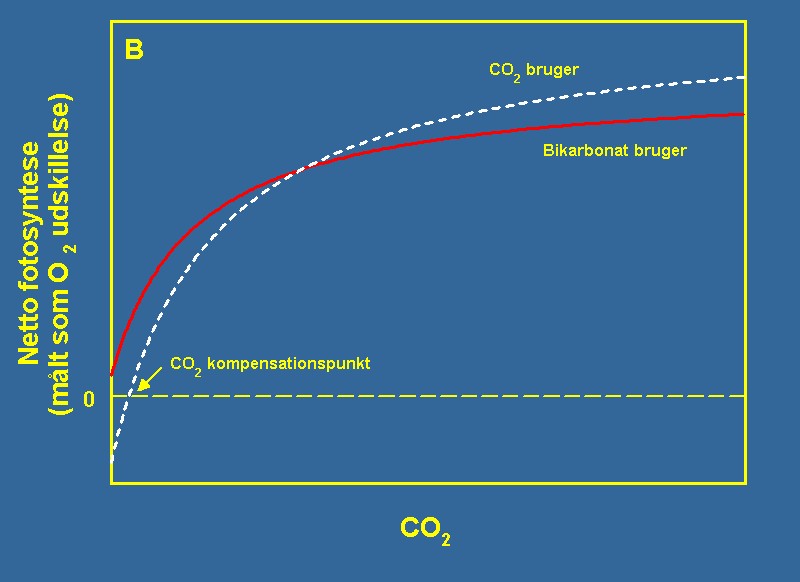
| Figur 2A og 2B Figuren viser den teoretiske sammenheng mellom lys og fotosyntese (A) og CO2 og fotosyntese (B). I begge situasjoner beskriver en metningsfunksjon sammenhengen selv om den aktuelle form på funksjonen er forskjellig. | |||
| |||
| For begge ressurser er kompensasjonspunktet definert ved det nivå hvor nettofotosyntesen eller veksten er lik null. Ved nivåer under kompensasjonspunktene kan planten ikke opprettholde sin biomasse. Ved en metningskurve er der likeledes et punkt, hvor ytterligere økning av ressursen ikke resulterer i økt fotosyntese eller vekst. Se dessuten teksten for ytterligere forklaring. | |||
| |||
| Interaksjon mellom lys og CO2 fremmer veksten hos vannplanter - del 2 | |||
| Vannplanter har som regel adgang til to former for uorganisk kullstoff: Karbondioksid (CO2) og bikarbonat (HCO3-). Hovedparten av vannplantene foretrekker CO2 fremfor HCO3-, da det opptas uten energiforbruk fra mediet og dessuten kan mange vannplanter ikke utnytte HCO3- direkte i fotosyntesen. Den generelle CO2-kinetik er vist på figur 2B. Formen på kurven avviker en smule fra lysutnyttelseskurven, da den ikke er lineær ved lave CO2-konsentrasjoner. Men også her kan vi fastlegge et CO2-kompensasjonspunkt som den konsentrasjon av CO2, hvor lavere konsentrasjoner resulterer i en negativ fotosyntese, og høyere konsentrasjoner i en positiv fotosyntese. Den ene av de to viste kurver har et CO2-kompensasjonspunkt på 0 mg CO2 per liter. Denne kurven illustrerer en HCO3--bruker, som klarer å fortsette fotosyntesen selv ved null CO2, da den kan bruke bikarbonat som kullstoffkilde i fotosyntesen. | |||
| I naturen er konsentrasjonen av CO2 i vannet ofte større enn i luften, men på tross av det, er den aktuelle tilgjengelighet av CO2 for vannplanten ofte lavere. Dette skyldes den langsomme diffusjon (molekylenes egenbevegelse) av gasser i vann, hvor diffusjonshastigheten er 10.000 gange lavere enn i luft. Selv om CO2-koncentrationen i mange vannløp er langt høyere enn i luften, resulterer den langsomme gassbevegelse i CO2-begrensning for veksten hos vannplantene. De tynne bladene, som typisk kjennetegner undervannsplanter, reduserer i høy grad CO2-begrensningen. Det skyldes dels at tynnere blader har tynnere grenselag (lag av stillestående vann) som CO2 skal diffundere gjennom og dels at det er kortere diffusjonsvei for CO2, når det endelig er inne i bladet og fikseres til karbohydrater i fotosyntesen. Det er formodentlig minst like vesentlig, at plantene er i stand til å opp- og nedregulere de forskjellige puljer av enzymer (eksempelvis RuBisCO og PEPcarboxylase), som deltager i kullstoffikseringen. Ved lave CO2-konsentrasjoner kan planten investere mer energi i enzymer, som hjelper i prosessen ved CO2-optakelsen og -fikseringen og dermed lindre effekten av CO2-begrensning. Visse planter kan dessuten produsere isoenzymer, som er enzymer med forskjellige kjemiske optima, hvor affiniteten for CO2 endres til fordel for mer effektivt CO2-optak. Det er fortsatt uvisst, hvor viktig rolle isoenzymerne spiller for CO2-opptaket og i den vitenskapelige litteratur er isoenzymer ofte forbundet med temperaturakklimatisering i planter. | |||
| |||
| De økologiske implikasjoner for interaksjonen mellom lys og CO2 er iøynefallende. Hvis foreksempel en økt CO2-konsentrasjon fremmer lysutnyttelseseffektiviteten, gir det vannplanterne mulighet for å søke dypere ned i vannsøylen, hvor lyset blir mindre, mens CO2-konsentrasjonen økes som følge av mineralisering av organisk stoff på sedimentoverflaten. Høy lystilgjengelighet kan på den anden side gi planten mulighet for å senke sitt CO2-kompensasjonspunkt (Maberly 1983, Maberly 1985). Det er spesielt en fordel for mattedannende planter som foreksempel mikroalger på steinoverflater. I slike systemer er der oftest rikelig med lys, men ekstremt lave CO2-konsentrasjoner i selve matten som følge av høy fotosyntese og liten vannutskiftning. Her spiller interaksjonen mellom lys og CO2 en stor rolle, da den høye lystilgjengelighet drastisk kan øke effektiviteten av CO2-ekstraktsjonen. | |||
| Interaksjon mellom lys og CO2 fremmer veksten hos vannplanter - del 3 | |||
|
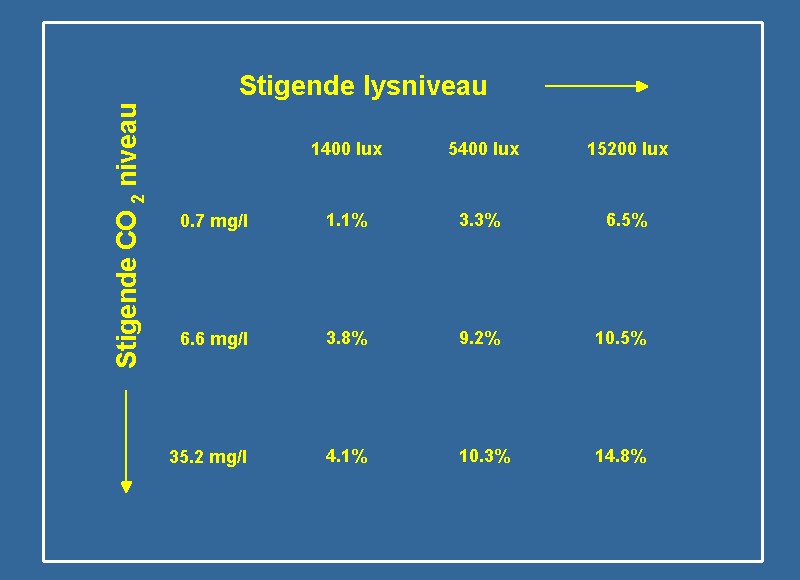
Akklimatisering til forhold med ressursbegrensninger er ofte en dyr affære for plantene. Enten det er en økt investering i klorofyll eller enzymer, så resulterer akklimatiseringen i et høyere behov for næringsstoffer og større energiforbruk. Det økede energiforbruket stammer fra proteinene, hvis funksjonalitet konstant skal opprettholdes for at cellen er funksjonsdyktig. Opprettholdelsen forbruker mye energi og karbohydrat, som ellers kunne være brukt til vekstformål. Det er derfor ikke alt det uorganiske kullstoff, som assimileres til karbohydrater, som går til vekstformål. Vi var interessert i å bestemme hvilken faktor - lys eller CO2 - som var den viktigste begrensende faktor for veksten hos vannplanter. Likeledes ønsket vi å vurdere, hvorvidt lys eller CO2 kunne vikariere for hverandre og dermed øke veksten ved foreksempel å tilføre mer lys ved sterk CO2-begrensning eller omvendt. For at belyse disse spørsmål eksperimentelt, dyrket vi Riccia fluitans i et matriksdesign ved mettede næringsstoffkonsentrasjoner (med næring menes det nitrogen og fosfor samt alle mikronæringsstofferne). Eksperimentet ble designet med henblikk på å regulere lys og CO2-konsentrasjonen uavhengig. Tabell 1 viser det eksperimentelle matriks, hvor vi hadde 9 behandlinger, som kun varierte ved forskjellig lysintensitet og CO2-konsentrasjoner. Den laveste lysintensitet svarer til forholdene tett på dybdegrensen for vannplanters vekst samt til lystilgengeligheten over et svakt belyst standardakvarium. Den høye lysintensitet svarer derimot til lysintensitet som forekommer i naturen tett på vannoverflaten eller i ekstremt velbelyste akvarier med høytrykks kvikksølvlamper eller halogenlamper. Den mellomste lysintensitet i matriksen er den intensitet, som svarer best til et velbelyst standardakvarium. Det lave CO2-nivået svarer til den CO2-konsentrasjon, som forekommer i de fleste sjøer, eller i akvarier uten CO2-berikning og eventuelt med en luftpumpe tilsluttet. Den høye CO2-konsentrasjonen er 40 mg/l, som hyppig forekommer i vannløp med grunnvannstilførsel - som er lik den maksimale CO2-konsentrasjonen, som anbefales av de mer erfarne planteakvarister. Vi brukte Riccia fluitans som en modellplante i våre eksperimenter, da den er lett å lage vekstmålinger på. Vi kunne derfor forholdsvis lett arbeide med mange replikaer, hvilket gjør, at vi kan tillate oss å dra sterkere konklusjoner fra våre eksperimenter. | |||
|
Tabell 1 Det eksperimentelle matriks viser det eksperimentelle design og de resulterende vekstrater som prosent per dag. Gående horisontalt fra venstre mod høyre økes lysintensiteten og gående vertikalt fra toppen mot bunnen økes CO2-konsentrasjonen. Lavt lys svarer til situasjonen i mange standardakvarier, mens lav CO2 svarer til et akvarium med luftpumpe (luftlikevekt). Til sammenligning er lysintensiteten ved høyt solskinn i det nordlige Europa ca. 7000 lux og en naturlig bekk kan inneholde opp til 50 mg CO2 per liter. Både lux og mg/l er foreldede enheter for lys og CO2, og disse enheter forekommer ikke i den vitenskapelige litteratur. Ved omregning av vitenskabelige enheter til disse generelle enheter har vi benyttet følgende faktorer: for lys (400 til 700 nm) svarer 1 µmol fotoner m-2 s-1 til 60,6 lux og for CO2 svarer 1mmol l-1 til 44 mg/l. | |||
| |||
|
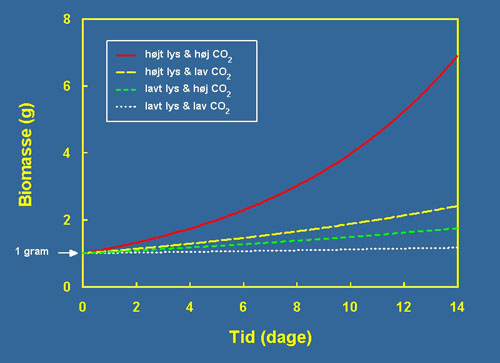
Tabell 1 viser resultatene fra våre eksperimenter. Vekstraten for Riccia fluitans er uttrykt i prosent per dag, hvor vi forutsetter eksponensiell(1) vekst i forsøksperioden. Det ses, at ved lavt lys og lavt CO2 kan Riccia fluitans akkurat opprettholde en positiv vekstrate, mens vekstraten ved lav CO2 og mye lys er nesten 6 gange større. Kanskje er det langt mer overraskende, at plantens vekst ved lavt lysnivå økes 4 gange ved tilsetning av CO2 til vannet! En 4 til 6 ganger økning av vekstraten virker kanskje ikke mye, men på grunn av vekstratens eksponensielle natur, skaper det en stor forskjell over en periode på eksempelvis to uker. Figur 3 viser, hvorledes 1 gram Riccia fluitans utvikler seg over to uker ved fire forskjellige vekstrater. Lavt lys og lav CO2-konsentrasjon resulterer knapt nok i positiv tilvekst over de to ukene, hvor behandlinger med mye CO2 og lav lysintensitet nærmest resulterer i en fordobling av biomassen. Til sammenligning vokser 1 gram til 2,5 gram biomasse ved høy lysintensitet og lav CO2-konsentrasjon. Det sier seg selv, at utbyttet ved både å øke lyset og CO2-konsentrasjon overskygger effekten av å kun øke en av ressursene. Ved høyeste lysintensitet og CO2-konsentrasjon, vil 1 gram Riccia fluitans vokse til overraskende 6,9 gram etter bare to uker. Det overraskende er, at vekstforøkelsen ved både å øke lys og CO2 samtidig er større enn bidraget fra hver av de to ressurser. Eksempel: Vekstraten ved lite lys og lite CO2 er 1,1% per dag. Ved stigende lysmengde vokser planten 3,3 % per dag eller et pluss på 2,2 % sammenlignet med startbetingelsen. Tilsvarende, ved å øke CO2-konsentrasjon er vekstraten nå 3,8% per dag eller et pluss på 2,7%. En additiv sammenheng ville resultere i en vekstrate på 6,0% per dag (1,1 + 2,2 + 2,7) men den resulterende vekstrate fra kombinasjonen av lys og CO2 er 9,2% per dag, hvilket er signifikant høyere. Tendensen er den samme ved høyere nivå av lys og CO2 (se Tabell 1). | |||
| Figur 3 Figuren viser hvordan 1 gram Riccia fluitans utvikler sig over to uker under de gitte lys- og CO2-betingelser. Ved lavt lys og lav CO2-konsentrasjon kan Riccia fluitans akkurat opprettholde sin biomasse og 1 gram blir til 1,16 gram etter to uker (den hvite linjen). Ved lavt lys og høy CO2- konsentrasjon blir 1 gram til 1,75 gram (den grønne linien) og ved høyt lys samt lav CO2- konsentrasjon blir 1 gram til 2,41 gram (den gule linjen). Kombinasjonen overstiger effekten fra de enkelte ressurser og ved høyt lys og høy CO2- konsentrasjon blir 1 gram til 6,90 gram (den røde linje). | |||
| |||
| (1) Ved forutsetning av eksponensiell vekst kan vi bruke følgende formel til beregning av vekstraten: µ = (ln W1 - ln W0)/t, hvor W0 er vekten av plantevevet ved forsøkets start, W1 er vekten etter inkubasjonen og t er inkubasjonstiden i dager. Bakgrunnen for at anta eksponensiell vekst er, at alt nytt vev som dannes under inkubasjonen i prinsippet kan ta del i veksten og skape mer nytt vev, og derfor blir det til en slags renters rente. | |||
| Interaksjon mellom lys og CO2 fremmer veksten hos vannplanter - del 4 | |||
| Figur 4 er en skjematisk og konseptuel illustrering av våre resultater. Ved lavt lys og lav CO2 er der ikke mye energi til rådighet for opp- og nedregulering av klorofyll- og enzympuljer. Hvis vi tilfører en smule mer CO2 til systemet reduseres plantens behov for energi til CO2-opptak, og dermed er det mer energi til optimering av lysutnyttelsen - mer klorofyll kan produseres uten fatale konsekvenser for energibudsjettet. Planten kan nå, uten at vi har økt lysintensiteten, utnytte den tilgjengelige lysmengde mer effektivt. Presis samme forklaring kan brukes til å forklare, hvorfor økt lys stimulerer veksten selv ved meget lave CO2-konsentrasjoner. Når der er mer lys til rådighet, er behovet for investering i lysutnyttelsessystemer mindre og den frie energi kan investeres i økt CO2-optag, således at den CO2, som er tilstede i vannet, kan ekstraheres med større effektivitet. | |||
| Figur 4 Forklaringsdiagrammet viser, hvordan den eksterne ressursmengde påvirker de interne utgifter i cellene til lysutnyttelse og CO2-assimilering. Fremstilt på denne simple illustrative måte, er det balansen mellom input og utgifter som bestemmer output i form av fotosyntese eller vekst. | |||
 Figur4 | |||
|
Våre resultater av vekstforsøk med Riccia fluitans kan forhåpentligvis overføres til de fleste vannplanter, og det siste tiåret har likeledes brakt vitenskabelig evidens, som underbygger denne påstand. Det har blitt utført eksperimenter med Elodea canadensis og Callitriche cochocarpa, som viser den samme tendens (se litteraturlisten), og det betyr, at ressursbegrensning ikke kan forklares med Liebigs simple baljemodell. Mange ressurser kan formodentlig vikariere for hverandre eller i det minste lindre symptomene på vekstbegrensning ved forskjellige grader av ressursbegrensning. Ved å se våre resultater i et litt større aspekt, kan vi forvente, at en stigning i atmosfærens CO2 vil resultere i en økt planteproduksjon på jorden. Vi kan dessuten forutse, at visse planter vil kunne fortynde deres puljer av kullstoffikserende enzymer, hvilket vil senke næringsverdien i korn, da enzymer er proteiner. En fordobling av CO2 i atmosfæren vil sannsynligvis ikke have noen effekt på produksjonen hos akvatiske planter, da mange planter allerede gror ved overmettede CO2-konsentrasjoner. For de få planter, som ikke gjør det, slike som undervannsplanter i sjøer, er det vanskelig å forutsi betydningen av økt CO2-tilgengelighet fordi vi her har konkurransen fra søfytoplanktonet og det kompliserende element av bikarbonatutnyttelse. | |||
| Hvordan kan vi så bruke informasjonen i hobbyplanteakvariet! På mange måter ligner det moderne planteakvarium våre eksperimentelle vekstmatriks med Riccia fluitans. Selv om det er vanskelig å kontrollere tilgengeligheten av hver enkel ressurs, kan vi definere den ønskede mengde av lys og CO2 samt hvor meget næring i form av nitrogen, fosfor, jern og mikronæringsstoffer, som vi vil tilby våre planter. Starter vi med næringsstoffene, vil et gjennomsnittlig planteakvarium med en anstendig fiskepopulasjon normalt ha rikelig med nitrogen og fosfor. Når det gjelder jern, kalium, natrium, mangan og andre mikronæringsstoffer, er det ofte mer komplisert. Noen akvarier er planlagt fra starten med eksempelvis lateritt og annen gjødende materiale i sedimentet, mens andre ikke er. For det meste kan en akvariums-plantegjødning uten nitrogen og fosfor trygt tilføres for opprettholdelse av sunn vekst. Det er ofte langt vanskeligere og også dyrere, når akvariet skal forsynes med intensivt lys. Både fluorescerende lys og høytrykks-kvikksølvlamper kan produsere det ønskede lys, hvis de forsynes med effektive reflektorer, men i dype akvarier (over 50 cm) er det vanskelig å oppnå nok lys til små lyskrevende planter i forgrunnen av akvariet. Med bakgrunn i våre eksperimenter, foreslår vi derfor en beriking med CO2 som viktigste utgangspunkt! Vi tror, at selv ved beskjeden lysintensitet, vil man oppleve en tydelig forbedring i plantenes vekst i akvariet. Den presise konsentrasjon av CO2 kan alltid diskuteres, men med mindre, at man har meget følsomme fisk i akvariet, vil konsentrasjoner fra 25 mg/l og op til 50 mg/l kun ha en stimulerende effekt. Man vil nå oppleve at planter, som før knapt kunne overleve, trives ved berikingen av CO2. | |||
|
Litteratur Andersen (1999) Interaktioner i mellem lys og CO2 fremmer væksten hos Riccia fluitans L. Rapport fra Ferskvandsbiologisk Laboratorium, Københavns Universitet. E-mail tandersen@zi.ku.dk Maberly (1983) The independence of photon irradiance and free carbon dioxide or bicarbonate concentrations on the photosynthesis compensation points of freshwater plants. New Phytologist 93: 1-12. Maberly (1985) Photosynthesis by Fontinalis antipyretica. I. Interaction between photon irradiance, concentration of carbon dioxide and temperature. New Phytologist 100: 127-140. Madsen (1993) Growth and photosynthetic acclimation by Ranunculus aquatilis L. in response to inorganic carbon availability. New Phytologist 125: 707-715. Madsen og Sand-Jensen (1994) The interactive effect of light and carbon on aquatic plant growth. Plant, Cell and Environment 17: 955-962. | |||

© www.tropica.com
Oversatt for AkvaForum av Geir Kristiansen

