Selvbygging av akvarium
Her er en tabell for glasstykkelser (cm). Dybde vertikalt, lengde horisontalt:
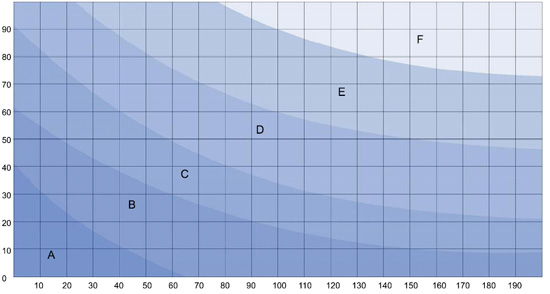
| SIDERUTER | BUNNPLATE | |
| A | 4 mm | 6 mm |
| B | 5 mm | 6 mm |
| C | 6 mm | 10 mm |
| D | 10 mm | 12 mm |
| E | 12 mm | 15 mm |
| F | 15 mm | 20 mm |
Eksempel 1, 112 liter enkelt akvarium
Vi trenger 4 vegger og bunn.
Størrelse 40cm x 40 cm x 70 cm = gruppe C = 6mm sider og 10mm bunn. Dette gir 112 liter brutto volum.
Netto volum med 4 cm fra kant til vannet gir: 35cm x 38,8cm x 68,8cm = 93,4 liter.
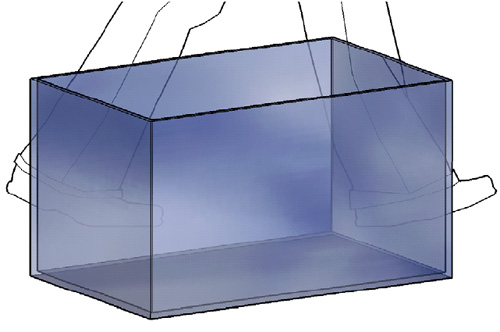
Mål:
(Forutsetter at silikonlimet bygger 2mm)
Bunn = D:40cm x L:70cm (10mm glass)
Langsider = H:38,8 (39)cm x L:70cm (6mm glass)
Kortsider = H:38,8 (39) x D:38,4cm (6mm glass)
H=høyde, L=lengde og D=dybde.
Eksempel 2, 126 liter akvarium med stort filterkammer
Vi trenger 4 vegger, bunn og en indre vegg med hull+utsparing til vann inn og ut.
Størrelse 45cm x 40cm x 70 cm = gruppe D = 10mm sider og 12mm bunn.
Dette gir 126 liter brutto volum. Netto volum med 4 cm fra kant ned til vann
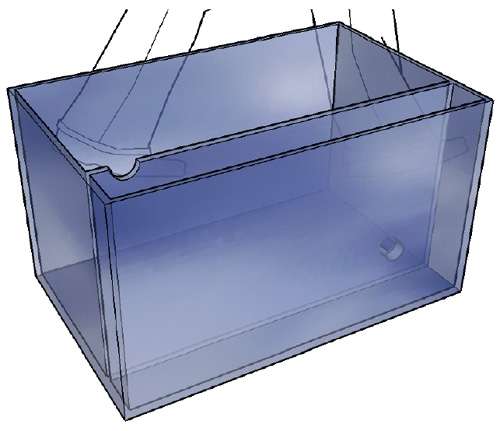
Akvariedel: 35,2cm x 68,8cm x 35cm = 84,8 liter.
Filterdel: 8,8cm x 68,8cm x 35cm = 21,2 liter.
Totalt 106ltr.
Mål:
(Forutsetter at silikonlimet bygger 2mm)
Bunn = D:45cm x L:70cm (10mm glass)
Langsider = H:39cm x L:70cm (6mm glass)
Kortsider = H:38,4cm x D:39cm (6mm glass)
Filtervegg = H:39cm x L:68,4 (6mm glass)
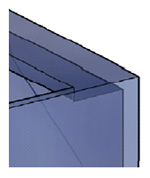 Toppdekselstøtte
Toppdekselstøtte
Om du ønsker kan du få laget en "hylle" langs hver kortside. Denne kan for eksempel være 2-4 cm bred og må være 39cm lang (= kortsiden) og limes inn flatt langs øverste kant som vist til høyre. For akvarium 2 blir lengden = avstanden fra frontglass til innervegg, en avstand kan du tilpasse selv så du får plass til den typen filter du vil ha. En blåfiltermatte kan skjæres til så det passer nesten uansett hvilken avstand du velger.
Toppdeksel
Til akvarium 1: 2 stk L:70 x D:20 (4-6mm glass),
1 hjørne bør kuttes av på begge for ledninger og fôring.
Til akvarium 2: 1 stk L:70 x D:20 + 1 stk L:70 x D:25, alternativt 2 stk L:70 x D:22,5.
Igjen med et hjørne på hver kuttet av for ledninger og fôring.
Vekt på akvariene
Glass har egenvekt på rundt 2,5 (litt avhenging av glasstype).
Akvariet i eksempel 1 har 7.847 cm3 glass eller 7,85 liter og veier da 19,6 kg + silikon.
Akvariet i eksempel 2 har litt mer glass, 9.823 cm3 eller 9,8 liter, og veier 24,5 kg + silikon.
Toppdeksler kommer i tillegg.
Liming av akvariet
Til limingen trenger du rødsprit, fyrstikker, tape og silikon (sort eller blank). Du kan bruke akvariesilikon fra en akvariebutikk, eller du kan bruke silikontuber fra nærmeste byggvarebutikk. Om du gjør det siste skal den være uten soppmiddel!
1. Alle kanter som skal limes må vaskes med rødsprit (avfetting) for å sikre god heft.
2. Den enkleste måten å bygge på er å bruke fyrstikker som avstandsklosser.
3. Bruk tape for å holde det hele sammen og bruk punkter av silikon (ca 5 på langside og 3 på de andre) og la stå et døgn.
4. Fjern fyrstikkene og legg på silikon fra innsiden. Vær omhyggelig på at silikonen
fyller helt 100% mellom glassene + en 45° kant langs hele fugen. Er du nøye med
pene kanter kan en strime tape (anbefaler 3M Scotch Magic tape) legges på hver side i
passe avstand, og du kan dra efter med en finger før du fjerner tapen, og så dra efter
(mykt og forsiktig!) med fingeren igjen, denne gang dyppet i sterkt Zalo-vann. (fordrer
at akvariet skylles nøye før bruk).
5. Om du bygger med skillevegg anbefales at denne settes inn til sist! Der kan være
vanskelig å komme til med silikonen ellers. En liten tube kan være greit når veggen
settes inn da det er trangt i filter-delen.
6. Det blir uvergelig silikonrester som tyter ut mellom glassene. Det gjør ingen ting,
heller tvert i mot! Det SKAL tyte i alle fall litt. La det være til det er tørt og fjern med
en skarp tapetkniv som holdes langsmed glasset. IKKE stikk kniven inn mellom
glassene, ikke engang litt!
7. La stå i minst 1 døgn, helst 2.
8. Test om akvariet er vanntett ved å fylle det og la stå med vann i et døgn før du tar det i
bruk. Om du finner utettheter så skal det hele være HELT TØRT før du reparerer.
Tips om plassering
Når akvariet er ferdig er det en fordel om du legger et underlag, for eksempel en 3mm Depronplate under. Dette vil ta opp ujevnheter og vil hindre for eksempel et sandkorn i å skrape opp eller påvirke brudd. Depronplater fås på byggvarebutikker.
Du kan også bruke en slik nederst i akvariet for å gi bunnglasset en beskyttelse mot at steiner og røtter med sand skaper spenninger mot glasser. Depron er også meget isolerende. For mye Depron rundt akvariet (bakvegg/sidevegg(er)) kan gi problemer med for varmt vann om sommeren.
Kilder:
Diverse Internett steder, inn- og utland herunder Akvaforum.
Design av akvarier av Helge Hafstad, utført i Google SketchUp
Tabell laget i Corel Draw 12 med basis i tabell fra Den Store Akvarieboken (Wennergren Cappelen)
Helge A. Hafstad - 02.11.2006

